

 联系我们
联系我们
最新研究,浙江大学研究团队在 Molecular Plant 发表的了题为“BIK1-mediated phosphorylation and SCF⁵ᴷᴵᴾ³¹-mediated ubiquitination coordinately control CNGC3 homeostasis and activity to fine-tune plant immunity”的研究论文。该研究揭示了拟南芥中 BIK1 激酶与 SCF⁵ᴷᴵᴾ³¹ 泛素连接酶组成的调控模块,通过协同调控 CNGC3 的磷酸化与泛素化,实现对Ca2+通道活性和蛋白稳态的精准控制,进而平衡植物生长与免疫应答,为作物抗逆育种提供了新的分子靶点。
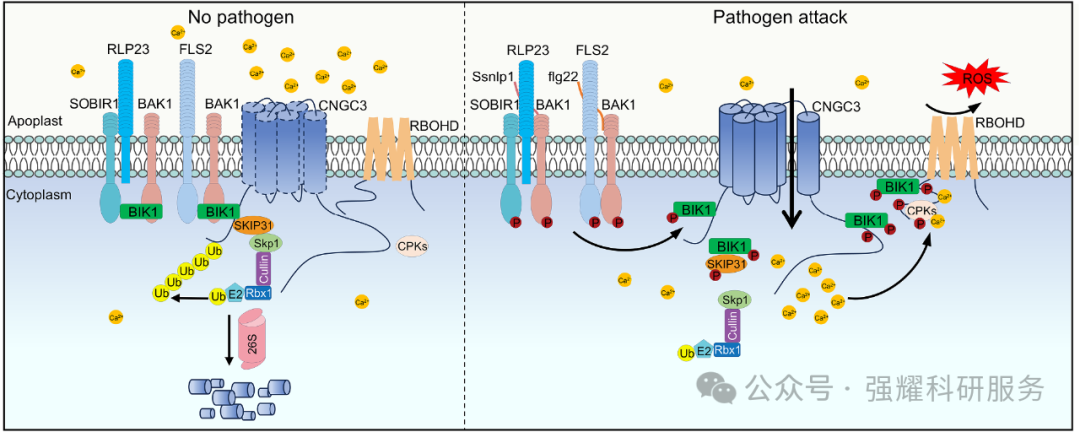
Fig. BIK1-SKIP31工作模型图
一、研究背景
植物通过两层先天免疫系统抵御病原菌侵袭:病原相关分子模式触发的免疫(PTI)和效应因子触发的免疫(ETI)。PTI 激活后会快速引发Ca2+内流、活性氧爆发等防御反应,其中Ca2+作为关键第二信使,其稳态调控对植物免疫至关重要。环核苷酸门控离子通道(CNGCs)是植物Ca2+内流的核心通道家族,参与生长发育、胁迫响应及免疫调控。已有研究表明部分 CNGC 成员的通道活性受磷酸化调控,但泛素化修饰在 CNGC 稳态调控中的作用及是否存在普适性调控机制仍不明确。核盘菌是一种破坏性极强的坏死营养型真菌,寄主范围广泛,目前缺乏高抗作物品种,亟需解析植物对其抗性的分子机制。
二、研究痛点
1. CNGC 家族的稳态调控机制(尤其是泛素化修饰)尚未明确,无法解释植物如何在生长与免疫间维持平衡。
2. 现有研究多聚焦单个 CNGC 成员的独立调控,缺乏对家族整体调控模式的探索。
3. 核盘菌抗性的分子机制仍需深入挖掘,亟需鉴定关键调控基因用于作物育种改良。
三、关键结果
1. CNGC3 是正向调控植物抗核盘菌的功能性Ca2+通道
突变体分析显示,cngc3 突变体接种核盘菌后病斑面积显著增大、病原菌生物量增加,而 CNGC3 过表达株系(CNGC3-OE)抗病性增强(图 1A-C)。CNGC3 定位于细胞质膜,核盘菌接种后 12 小时表达量显著诱导(图 1F);钙成像和膜片钳实验证实,CNGC3 可介导 PAMP 诱导的Ca2+内流,其 P 环区域 G361 位点突变(CNGC3ᴳ³⁶¹ᴱ)会丧失通道活性(图 1L-Q)。cngc3 突变体中 PTI 相关基因(WRKY29、FRK1、PR1、PR5)表达下调,ROS 爆发和Ca2+内流减弱,而 CNGC3-OE 株系则相反(图 1D-K)。
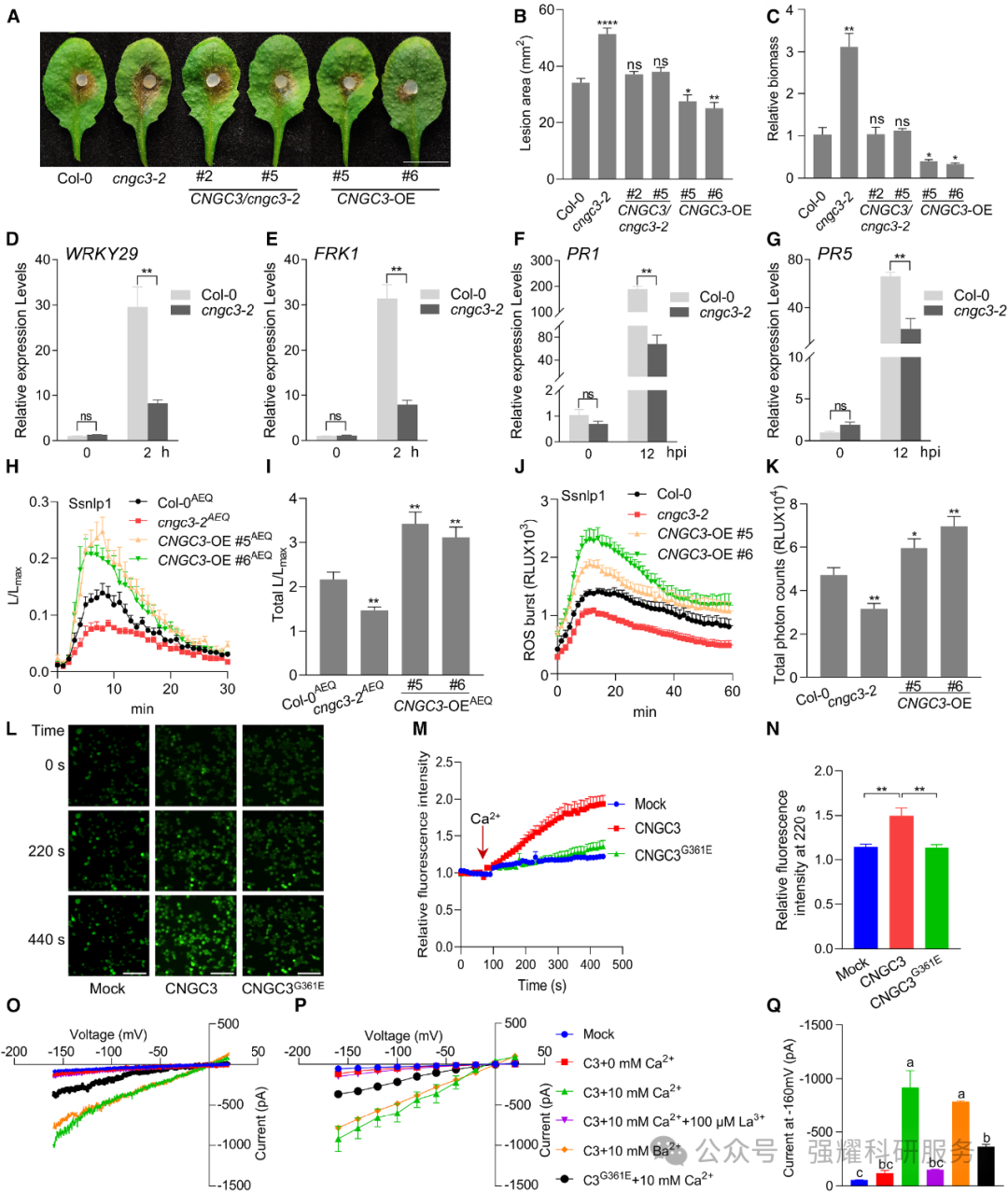
Fig1. CNGC3 是一种Ca2+渗透通道并促进植物对菌核的抗性
2. BIK1 通过磷酸化 CNGC3 增强其通道活性
体外激酶实验和体内磷酸化检测证实,BIK1 可与 CNGC3 直接相互作用并磷酸化其胞质 C 端的 4 个 Ser 位点(S537、S596、S631、S668)(图 2A-F)。
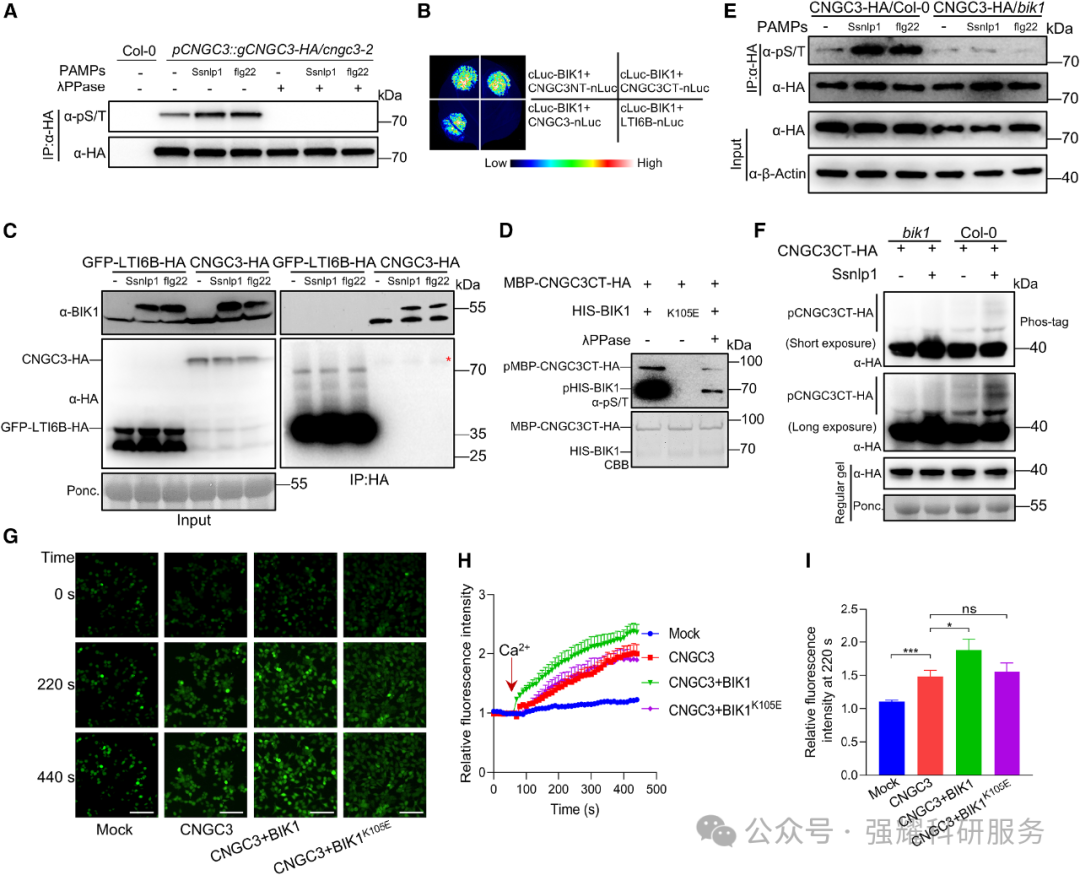
Fig2. BIK1 与 CNGC3 的相互作用及磷酸化修饰验证
钙成像实验显示,BIK1 可显著增强 CNGC3 的Ca2+通道活性,而激酶失活型 BIK1ᴷ¹⁰⁵ᴱ无此效应;CNGC3 的 4 个 Ser 位点突变为 Ala(CNGC3⁴ᴬ)后通道活性丧失,突变为 Asp(CNGC3⁴ᴰ)后通道活性增强(图 2G-I、图 3D-F)。抗病性实验表明,CNGC3⁴ᴰ互补株系对核盘菌的抗性强于野生型互补株系,而 CNGC3⁴ᴬ互补株系无法恢复 cngc3 突变体的感病表型(图 3H-J)。
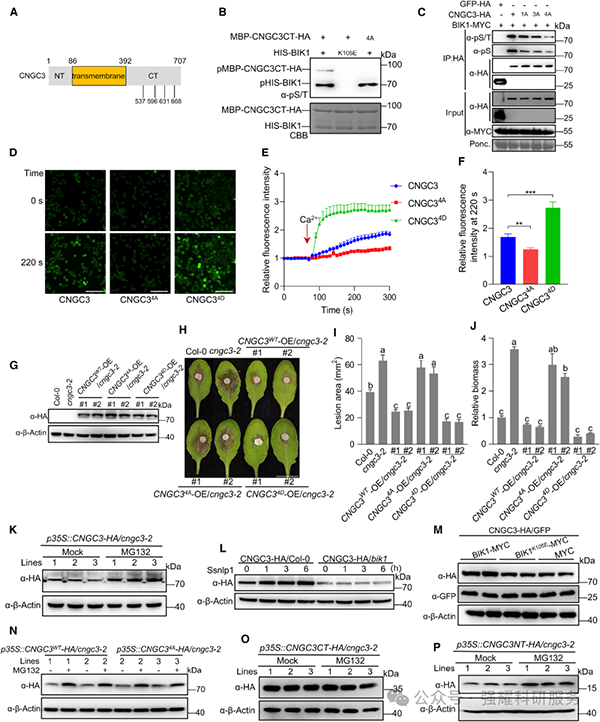
Fig3. BIK1 磷酸化 CNGC3 的位点功能及 CNGC3 的降解机制
3. SKIP31 通过泛素化降解 CNGC3 负调控植物免疫
Y2H 和 Co-IP 实验证实,SKIP31 与 CNGC3 的 N 端相互作用,且该相互作用可被 PAMP 处理抑制(图 4A-F)。泛素化实验显示,SKIP31 可通过 K-X-V-R 基序中的 Lys8、Lys22 和 Lys33 位点泛素化 CNGC3,促进其通过 26S 蛋白酶体降解;skip31 突变体中 CNGC3 蛋白稳定性显著提高(图 4I-K、图 4M-N)。抗病性分析表明,skip31 突变体对核盘菌抗性增强,SKIP31 过表达株系感病性增加,且 cngc3/skip31 双突变体与 cngc3 单突变体表型一致,证明 SKIP31 通过调控 CNGC3 发挥作用(图 4O-Q)。
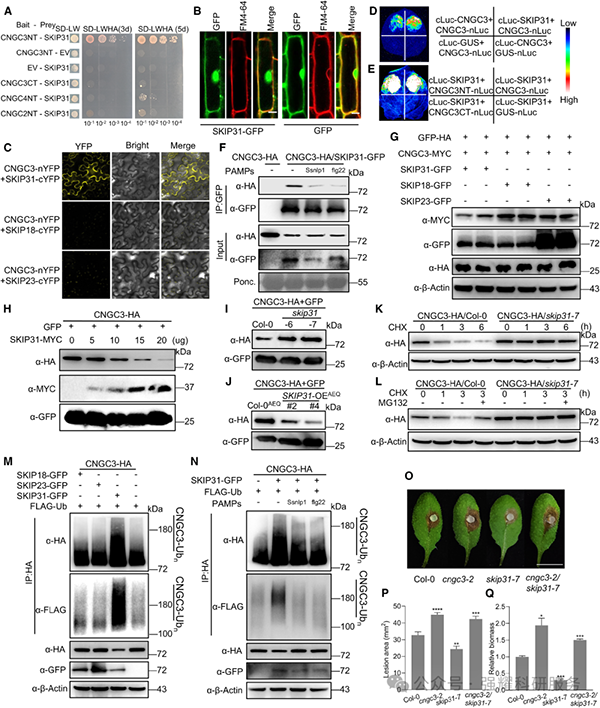
Fig4. SKIP31 与 CNGC3 的相互作用、泛素化及抗病功能
4. BIK1 磷酸化 SKIP31 抑制其泛素连接酶活性
BIK1 可与 SKIP31 直接相互作用并磷酸化其 5 个 Ser 位点(S88、S93、S125、S198、S261)(图 5A-H)。磷酸化位点突变实验显示,S88 和 S93 磷酸化可阻断 SKIP31 与 Skp1 的相互作用,S261 磷酸化可抑制 SKIP31 与 CNGC3 的结合,最终抑制 SKIP31 介导的 CNGC3 泛素化降解(图 5I-K)。抗病性实验表明,SKIP31 的 5 个 Ser 位点突变为 Asp(SKIP31⁵ᴰ)后无法互补 skip31 突变体的抗病表型,而突变为 Ala(SKIP31⁵ᴬ)可完全互补(图 5M-O)。
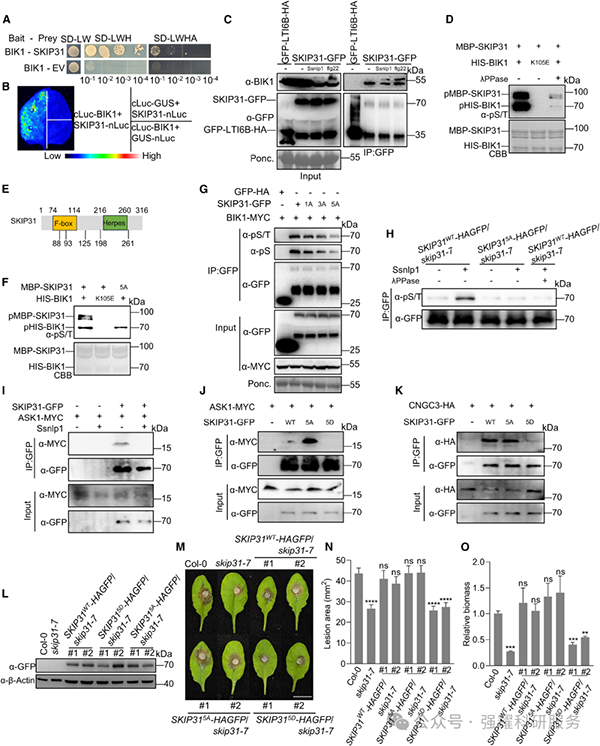
Fig5. BIK1 磷酸化 SKIP31 的位点及功能验证
5. 油菜中 BnCNGC3 和 BnSKIP31 功能保守
油菜 BnCNGC3ᴄ⁰⁴和 BnCNGC3ᴬ⁰⁵可互补拟南芥 cngc3 突变体的感病表型,其过表达株系对核盘菌抗性增强;Bncngc3 双突变体感病性增加(图 6A-F)。油菜 BnSKIP31 家族可与 BnCNGC3 相互作用并促进其泛素化降解;Bnskip31 四重敲除突变体对核盘菌的抗性显著增强,且生长发育无明显缺陷(图 6G-M)。
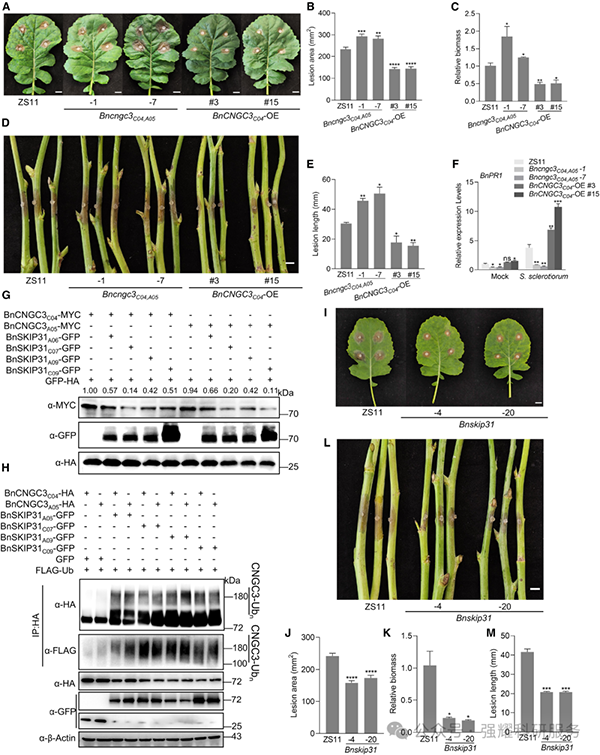
Fig6. 油菜中 BnCNGC3 和 BnSKIP31 的功能保守性
四、总结与展望
该研究阐明了 “BIK1-SKIP31-CNGC3” 调控模块的分子机制:无病原菌时,SKIP31 通过泛素化降解 CNGC3 抑制免疫反应,保障植物正常生长;病原菌入侵时,PAMP 激活 BIK1,一方面磷酸化 CNGC3 增强其Ca2+通道活性,另一方面磷酸化 SKIP31 抑制其泛素连接酶功能,稳定 CNGC3 蛋白积累,最终通过Ca2+信号放大 PTI 反应(图 7)。该机制在拟南芥和油菜中高度保守,揭示了磷酸化-泛素化协同调控 CNGC 稳态的普适性模式。研究还发现这些磷酸化位点在多种植物SKIP31中高度保守,且SKIP31能与所有测试的CNGC家族成员互作,暗示这是植物免疫调控的普遍机制。该工作首次阐明了SCF泛素连接酶与受体样激酶(RLCK)协同调控CNGCs的分子开关,为理解植物如何平衡生长与免疫提供了新视角。
原文链接:DOI:10.1016/j.molp.2025.09.012
本研究中SsNLP1多肽由强耀生物提供,强耀生物提供多肽合成,蛋白和抗体制备等优秀服务助力科研。
 返回
返回